
Efficacious Age Reversal in the Mouse Model with Subcutaneous Injections of Distinctive Cytokine Matrixes
Ron Shane, Ph.D., OMD, Andrei Golovko, Benjamin Morpurgo, Derek Chen, Karan Pandher, and Harumi Kimura
|
| |
Abstract
The intent of this study was to demonstrate that cytokine therapies from distinctive stem cell sources as shown in an earlier study could reverse cellular aging in a mammalian organism. It was shown by Shane 2012 et al that older human fibroblasts in culture when treated with peptide matrixes from human embryonic stem cells could express a gene profile characteristic of a younger phenotype. Dr. Rando’s work where he attached the blood supply of a younger and older mouse likewise showed that cytokines from a younger phenotype auspiciously modulated the transcriptional and translational machinery of the older mouse.
In this study we employed three distinctive cytokine matrixes from different stem cell sources. Moreover, there were different subcutaneous titration regimes which were utilized in this study as well as blending divergent peptide matrixes. The results from these cytokine interventions demonstrated that it is possible to ameliorate the gene expression profile of an older treated mouse phenotype where it becomes 70 percent identical to a younger untreated mouse and only 28 percent concordant with an older untreated control.
Thousands of genes were up and down regulated as a function of these distinctive cytokine configurations. Furthermore, hundreds of genes were likewise up and down regulated at least two fold. Pathologists from different universities did not find any indication of mutagenesis or neoplasia in tissues from bone, muscle, liver, heart, brain, and skin. A larger sample of mice will be employed in a future study for a longer duration than four months in order to comprehend if cytokine therapies can propitiously induce tissue remodeling in an older mammalian phenotype. These innovative regenerative protocols may be very auspicious to the older human phenotype. This study clearly shows that cytokine matrixes can reverse cellular quiescence in the mouse model where their transcriptional machinery is positively impacted to be concordant with a younger phenotype.
Introduction
Many research endeavors have attempted to employ stem cell sources either autologous or allogenic in order to temper cellular aging. These studies have been executed in a diverse array of animals and humans where stem cell protocols have been employed and are not very effective at reversing cellular aging. Many distinctive stem cell regimes have been utilized by medical researchers without any significant therapeutic outcome. Many studies involving the older phenotype where researchers have attempted to reverse cellular quiescence have been clinically irrelevant.
The use of stem cell strategies either IV as well as regional titration has not yielded medicinal results in terms of modulating gene changes associated with programmed aging. The heuristic benefits of stem cells themselves for mollifying the genetic changes involved with cellular aging is not effective in the mammalian phenotype. Systemic injections of stem cells have consistently demonstrated that these so called age attenuating strategies do not adhere to appropriate tissue sites for a prolonged period of time; and most importantly, systemic diffusion is not possible with stem cells. In general, stem cells are not an efficacious protocol for age attenuation. However, in our view, their paracrine peptides would be much more efficacious as an ameliorative molecular strategy for mitigating aging.
Shane 2012 et al showed that cytokine matrixes from embryonic stem cells did significantly change the gene expression profile of older human fibroblasts where they were more concordant with their daughters’ phenotype. These dermal fibroblasts were treated for 5 days in culture with cytokine configurations from human embryonic stem cells. The intent of our proposed proof of concept study is to determine if this prophylaxis is effective in a mammalian organism. There has been a bevy of studies in the preceding 15 years attempting to utilize a variety of stem cell sources both autologous and allogenic to reverse cellular aging in humans and other mammalian genotypes. These protocols were ineffective and oftentimes problematic.
We intend to utilize three distinctive cytokine matrixes as well as a diverse array of subcutaneous dosing regimes on the older mouse phenotype. This research paradigm has never been employed on the mammalian genotype. Cytokine matrixes to be used in this study will be derived from mouse embryonic stem cells as well as human umbilical cord lining progenitor cells. Moreover, a variety of peptides will be extracted from human placenta. All treated mice will be over 50 weeks old.
Dr. Rando’s work on mice involved “heterochronic parabiosis”. He demonstrated that cytokines of a younger mouse amelioratively impacted many physiological parameters of the older phenotype. These two organisms shared the same blood supply as they were surgically attached. Thus the older and younger mouse shared cytokines or other paracrine related signals. Dr. Rando observed that muscle as well as hepatic tissue was improved in the older phenotype, and also the older mouse’s wound healing response. Conversely, the younger mouse’s molecular robustness was tempered. Transcriptional and translational processes were affected by the succinct cytokine configuration titrating the receptors of various cell lines in these two surgically attached organisms. The focus of this preliminary study is to determine if cytokine configurations from undifferentiated stem cell sources can reverse the activities of genes in the older phenotype. We predict that the overall gene expression profile of the older treated mice can be propitiously remodeled characteristic of a younger phenotype. Cytokine prophylaxis has similar pharmakinetics to endocrine hormones in terms of how these peptides affect a diverse array of cells in the viable organism. Conversely, stem cells if delivered intravenously or through subcutaneous injections do not profusely circulate through the entire organism. Furthermore, we predict that the 200 proteins contained in our subcutaneous injections will not induce adverse reactions.
Muscle, cardiac, brain, hepatic, and skin tissues will be extracted from older and younger controls as well as the three groups of treated mice. Two pathologists will look at all the tissue specimens for any kind of tumor genesis or neoplasia. Food intake and normal nocturnal behaviors will be likewise observed in the treated groups and the various young and older untreated controls. It is predicted even at the highest dose of cytokine load these organisms will not have an adverse reaction. The physiological benefits of these cytokine therapies will not induce any kind of adverse response. We predict that the subcutaneous prophylaxis will essentially be propitious in terms of optimizing cellular transcriptional machinery.
Three groups of mice consisting of four organisms will receive a high dose of a distinctive cytokine configuration for four months. Group 1 will be titrated with cytokines from mouse embryonic stem cells; group 2 peptide matrixes derived from human placenta; and group 3 will receive a cytokine matrix extracted from stem cells harbored in human umbilical cord lining. In the second phase of this study two groups will be titrated four times a week for six weeks. Lastly, in the third facet of this study, group 1 will receive a cytokine configuration from human umbilical cord lining as well as peptides derived from human placenta. Conversely, in the group 2, these organisms will be titrated with peptides from embryonic stem cells and cytokine matrixes derived from umbilical cord lining. All treated organisms will receive four subcutaneous injections per week for a period of six weeks.
We are predicting that cytokine injections will temper over a period of time the progressive decline in these organisms’ physiology associated with aging as well as their tissue dysfunctionality. This unique therapy may be ameliorative to the genetic aberrancies involved with aging such as DNA fragmentation or breaks in chromatin as well as the intercellular devastation engendered by oxidative stress. Furthermore, we believe that titrating with cytokines from undifferentiated stem cell sources can induce overall tissue regeneration in many mammalian species. In our view, extraneous cytokine matrixes can auspiciously modulate compromised molecular pathways in the older phenotype so they become homeostatically robust.
Human aging is a multifactorial physiological phenomenon. The re-education of succinct genes which are dysregulated as a result of the molecular dynamics of aging will alleviate some of the deleterious physiological events engendered by biological aging. In our opinion, this proof of concept study should indicate that stem cells should not necessarily be utilized themselves for treating molecular aging; but instead the paracrine peptides are far more efficacious and can reverse the gene virulence associated with biological aging. It is predicted that the mammalian phenotype when treated with a cytokine regime will express a genetic profile characteristic of a younger phenotype.
MATERIALS AND METHODS
Cytokine matrixes.
Production of embryonic stem cell growth factors was performed according to the following protocol. Wild type ES cells in C57BL/6 background were cultured according to the standard TIGM ES cell expansion protocol for 3-4 days until the cultures are 70% confluent. At that time the media was collected and saved in 1 ml aliquots at -80oC and used later for injections. In order to maintain undifferentiated state of the culture, the cells were grown on inactivated MEF in presence of Leukemia Inhibitory Factor. About 200 ml of the MEF-conditioned medium were also collected and saved.
A pharmacological company from Singapore provided their Calecim product which consists of the cultural media or cytokine matrix derived from stem cells extracted from umbilical cord lining. Their cytokine matrix has been used in wound healing studies in humans; and is currently employed in a topical skin line.
Melsmon is a cytokine matrix derived from mesenchymal stem cells from human placenta. This peptide configuration is manufactured in Japan and has been used in Asia for over 60 years
Preparation of the working solutions.
The undiluted samples collected from plates with the ES cells represented “high concentration”; “moderate concentration” was achieved by 1:1 dilution of the samples with ES cell medium.
Dilutant was prepared by mixing 0.6 ml of benzyl alcohol and 39.4 ml of a saline solution.
Calecim was prepared by mixing 3 ml of Calecim stock and 12 ml of Dilutant
Melsmon was prepared by mixing of Melsmon stock and 12 ml of Dilutant
Animal procedures.
All animals were housed in the TIGM vivarium Research Core, and were handled in accordance with the principles and procedure of the Guide for the Care and Use of Laboratory Animals. All procedures were approved by The Texas A&M Institutional Animal Care and Use Committee (IACUC).
Experiment 1. Injections of the ES cell conditioned medium.
TIGM has procured twenty C57BL/6 males, 50 weeks of age, from The Jackson Labs. TIGM also had animals of the same genetic background of various age in the colony to be used as controls.
Beginning of the experiment tissue collection: muscle and skin samples were collected and frozen down immediately in RNAlater to preserve RNA prior to this study from the following animals:
- 3 aged mice (50 weeks of age)
- 3 young mice (10 weeks of age)
- 2 mid-age mice (36 weeks of age)
- 1 mid-age mouse (28 weeks of age)
Also, bone, brain, skin, muscle, heart and liver were collected from these animals and fixed in formalin for histological analysis.
The first treatment group consisted of 4 mice and they were provided with a “moderate” concentration of embryonic stem cell growth factors (100 ul), injected intravenously (IV) once a week for a total of 6 treatments followed by subcutaneous (sub-Q) injections of 150 ul for a total of 10 treatments for a four month duration. Four other older female mice were given a “high” concentration of embryonic stem cell growth factors for 16 weeks where they received an injection once a week as well (6 IV and 10 sub-Q injections). Lastly, in the third group, which is the control portion of the study, four animals were furnished with a Feeder conditioned medium injections (6 IV and 10 sub-Q injections). A group of 3 animals didn’t receive any treatments and were kept as controls. The mice were housed individually.
Experiment 2. Injections of Calecim and Melsmon.
TIGM provided 14 males of the outbred strain CD1 (ICR), 50-52 weeks of age as well as younger control males (32 weeks old) for the experiment.
Beginning of the experiment tissue collection: muscle and skin samples were collected and frozen down immediately in RNAlater to preserve RNA prior to this study from the following animals:
- 3 aged mice (50 weeks of age)
- 3 younger mice (32 weeks of age)
Also, bone, brain, skin, muscle, heart and liver were collected from these animals and fixed in formalin for histological analysis.
Each aged mouse was injected once every week 3 times x 50 ul subcutaneously: one dorsal, one ventral and one lateral (either left or right) during a period of 16 weeks. Location of the subsequent injection need to be shifted slightly by 0.5-1 cm moving along anterior-posterior axis. The mice were injected as follows:
4 aged animals received 3 x 50 ul of Dilutant during single session each week x 16 weeks
4 aged animals received 3 x 50 ul of Calecim during single session each week x 16 weeks
4 aged animals received 3 x 50 ul of Melsmon during single session each week x 16 weeks
1 younger animal received 3 x 50 ul of Melsmon during single session each week x 16 weeks
2 aged animals were kept as a control group, no injections.
The mice were housed individually.
Experiment 3. More frequent injections of Calecim and ES cell-conditioned medium over a short period of time.
TIGM provided 2 CD1 (ICR) males, 50-52 weeks of age. ES cell-conditioned medium, Calecim were prepared as described above.
Each mouse received 24 injection rounds over a period of 6 weeks (4 injections per week). During each injection round the mice were injected subcutaneously 3 times with 75 ul: one dorsal, one ventral and one lateral (either left or right). One mouse received ES cell-conditioned medium and the other was injected with Calecim.
The mice were housed individually.
Experiment 4. Combined injections of Calecim, Melsmon and ES cell-conditioned medium.
TIGM provided 2 CD1 (ICR) males, 50-52 weeks of age. ES cell-conditioned medium, Calecim were prepared as described above.
Each mouse received 24 injection rounds over a period of 6 weeks (4 injections per week). During each injection round the mice were injected subcutaneously 4 times with 50 ul: one dorsal, one ventral and 2 lateral (left and right). One mouse received 2 x 50 ul of Calecim and 2 x 50 ul of ES cell medium and the other 2 x 50 ul of Calecim and 2 x 50 ul of Melsmon.
The mice were housed individually.
End of experiment tissue and sample collection.
200-800 ul of blood serum were collected from each experimental animal via cardiac puncture. Muscle and skin samples were collected and frozen down immediately in RNAlater to preserve RNA. Also, bone, brain, skin, muscle, heart and liver were collected from these animals and fixed in formalin for histological analysis.
Histology
Formaldehyde-preserved tissues were fixed in paraffin, sectioned and stained with hematoxylin and eosin by Veterinary Pathobiology lab, Texas A&M College of Veterinary Medicine, College Station, TX.
Analysis of blood metabolites
ELISA kits to detect TNFa, IL1, IL6, IGF1, and Cortisol in mouse serum were purchased from Enzo Life Sciences, Farmingdale, NY, USA. Mouse Estradiol EIA kit was purchased from Cayman Chemical Company, Ann Arbor, MI. Serum samples were diluted and assayed according to manufacturer’s specifications. Plate reader capable of measuring absorbance at 405-420 nm was used for detection. Samples were run in duplicates.
Microarray analysis
RNA was isolated using RNAeasy Kit from Qiagen (Hilden, Germany) according to manufacturer’s specifications. Due to budgetary constraints, only one mouse was analyzed from each experimental group. Following quality assessment on an Agilent 2100 Bioanalyzer, 1 ug total RNA was used to generate biotin-labeled cRNA via a modified Eberwine RNA amplification protocol using the Ambion MessageAmp II-Biotin Enchanced kit (Ambion, Austin, TX). Labeled cRNA was applied to CodeLink mouse genome arrays (Applied Microarray, Tempe, AZ) for an 18 h hybridization followed by washing, staining, and scanning as per the CodeLink protocol. An Axon 4000B scanner was used for scanning the arrays. Array images were processed using CodeLink system software and global median normalization was used to generate normalized expression values. Microarray analysis was performed at the Department Nutrition and Food Science, Texas A&M University. Data analysis was performed using in-house algorithm to sort out meaningful changes and compare them between two groups. Also, cluster analysis was per formed using Statistica software package.
RESULTS
Analysis of blood metabolites
Estradiol.
ES cell medium was shown to increase estrogen levels, but the question remains regarding what was the reference levels (Figure 1). In the first experiment younger mice have lower estrogen levels that the older ones; in the other experiments it's the other way around, but it could be because one of the young mouse samples used in Experiments 2, 3 an 4 was significantly higher. Melsmon seems to lower estrogen a bit, Calecim lowers it too in the experiments 3 and 4 (4 injections a week), but increases it in the study 1 (single injection a week).
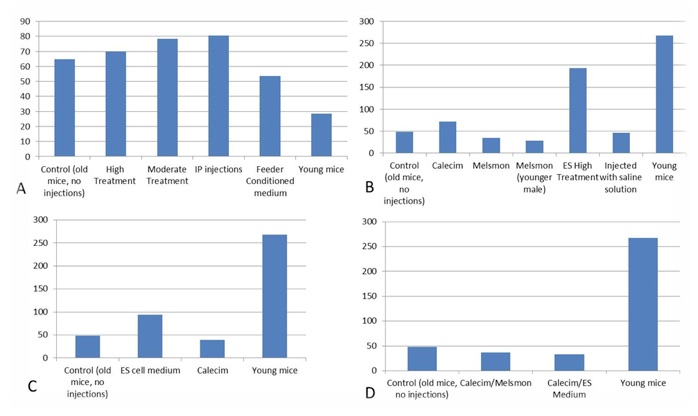
Figure 1. Estradiol, pg/mL A. 1st study: injection of ES cell medium; B. 2nd study: injection of Calecim and Melsmon; C. 3rd study: 4 injections per week x 6 weeks; D. 4th study: combined injections, 4 per week.
IGF1
Changes in the amount of IGF were very incremental, varied among different treatment groups within 5-10% (Figure 2). Experiments 2, 3 and 4 showed that IGF1 was slightly elevated in younger animals as compared to the old controls, and stand-alone Calecim and Melsmon treatments also slightly increased the levels if this metabolite. However, combined treatments with Calecim + Melsmon and Calecim + ES cell-conditioned medium lowered IGF1. Also high concentration of the ES cell-conditioned medium also lowered IGF1.
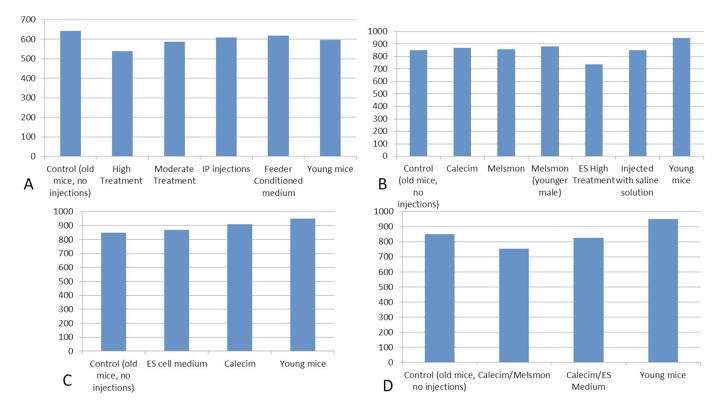
Figure 2. IGF1, ng/mL. A. 1st study: injection of ES cell medium; B. 2nd study: injection of Calecim and Melsmon; C. 3rd study: 4 injections per week x 6 weeks; D. 4th study: combined injections, 4 per week.
Cortisol.
All the samples were off the chart too high. This is consistent with mouse IgG in the samples interfering with the assay. It prevents the labeled cortisol from binding to the surface of the wells mimicking high non-labeled cortisol in the sample.
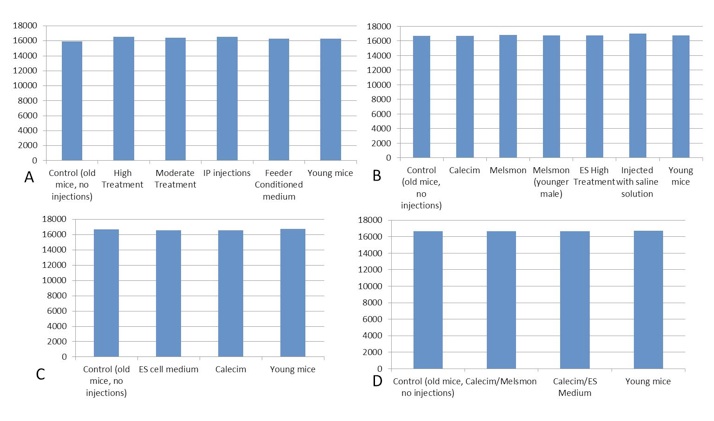
Figure 3. Cortisol, pg/ml. A. 1st study: injection of ES cell medium; B. 2nd study: injection of Calecim and Melsmon; C. 3rd study: 4 injections per week x 6 weeks; D. 4th study: combined injections, 4 per week.
IL1b, IL-6 and TNFa.
All samples had really low levels or non-detected levels at all. All the readings for the samples are in the noisy part of the curve that is barely above background levels of absorption. Apparently this is consistent with the normal state of these metabolites: if there is no inflammation going on, these metabolites may be at very low levels. If levels go into detectable levels, then there's some inflammation.
Microarray gene expression analysis
Data obtained in the form of Normalized Intensity was analyzed via two routes: fold change comparison between pairs of samples one of which included old and young mouse and the other included old vs. treated mouse. All our microarray findings are based in terms of changes on the comparison of the old untreated mouse with the overlap or similarities of the young untreated mouse with the old treated mice who were titrated with cytokine intervention. The other approach was cluster analysis.
Here’s the summary of the fold change comparison in the microarray data analysis.
Experiment 1. Injections of the ES cell conditioned medium.
Summary of results is shown in Table 1. The old treated mouse has over 60 percent of its genes similar to the younger untreated phenotype. Conversely, it has been observed that the treated older mouse and the younger untreated mouse is now 40 percent similar to the older untreated mouse. This connotes that the gene expression profile of the older treated mouse and younger untreated mouse sufficiently overlaps and are both 40 percent similar to the older untreated phenotype.
Experiment 2. Injections of Calecim and Melsmon.
Calecim
Summary of results is shown in Table 1. The old treated mouse has over 70% (skin) and 73% (muscle) of its genes similar to the younger untreated phenotype. Conversely, it has been observed that the treated older mouse and the younger untreated mouse is now 27% (muscle) and 30% (skin) similar to the older untreated mouse. This connotes that the gene expression profile of the older treated mouse and younger untreated mouse sufficiently overlaps and are both 27-30 percent similar to the older untreated phenotype.
Melsmon
The old treated mouse has over 69% (skin) and 72% (muscle) of its genes similar to the younger untreated phenotype. Conversely, it has been observed that the treated older mouse and the younger untreated mouse is now 28% (muscle) and 31% (skin) similar to the older untreated mouse. This connotes that the gene expression profile of the older treated mouse and younger untreated mouse sufficiently overlaps and are both 28-31 percent similar to the older untreated phenotype.
Experiment 3. More frequent injections of Calecim and ES cell-conditioned medium over a short period of time
Summary of results is shown in Table 1.
Overlap between No injection / young mouse change and No injection / ES medium treatment change in muscle: Up-regulated in both: 4967 genes, 39 more than 2-fold Down-regulated in both: 7121 genes, 215 more than 2-fold.
Overlap between No injection / young mouse change and No injection / Calecim treatment change in muscle: Up-regulated in both: 5839 genes, 69 more than 2-fold Down-regulated in both: 6968 genes, 241 more than 2-fold
We can conclude that Calecim is more potent than ES medium. But comparing the combined results from all studies (Table 1), it looks like the 4-months treatments with 1 injection/day produced slightly stronger effect than 4 injections/week for 6 weeks: more genes up- or down-regulated in general and more genes with 2-fold changes. Cluster analysis shows that the expression profiles look different than those of the young and old untreated mice.
Experiment 4. Combined injections of Calecim, Melsmon and ES cell-conditioned medium.
Overlap between No injection / Calecim+Melsmon treatment change and No injection / young mouse change in muscle: Up-regulated in both: 5180 genes, 69 more than 2-fold Down-regulated in both: 5933 genes, 355 more than 2-fold.
Overlap between No injection / Calecim+Melsmon treatment change and No injection / young mouse change in skin: Up-regulated in both: 4978 genes, 75 more than 2-fold Down-regulated in both: 7511 genes, 300 more than 2-fold.
Overlap between No injection / Calecim+ES treatment change and No injection / young mouse change in skin: Up-regulated in both: 4117 genes, 54 more than 2-fold Down-regulated in both: 6567 genes, 119 more than 2-fold.
We can conclude that Calecim+Melsmon is more potent than Calecim+ES medium: Calecim+Melsmon causes more gene expression changes similar to that of old -> young mouse than Calecim+ES. But if we compare the combined results from all studies (Table 1 below), it looks like the 4-months treatments with 1 Calecim injection/day produced slightly stronger effect than 4 plain Calecim or Calecim+Melsmon injections/week for 6 weeks: more genes up- or down-regulated in general and more genes with 2-fold changes.
| |
Genes up-regulated in both |
Up-regulated more than 2-fold in both |
Genes down-regulated in both |
Down-regulated more than 2-fold in both |
| ES cell medium |
|
|
|
|
| Muscle |
6949 |
98 |
5752 |
51 |
| ES cell medium 4 injections/wk |
|
|
|
|
| Muscle |
4967 |
39 |
7121 |
215 |
| ES cell medium |
|
|
|
|
| Skin |
7666 |
66 |
6201 |
54 |
| Calecim |
|
|
|
|
| Muscle |
9874 |
136 |
5536 |
327 |
| Calecim, 4 injections/wk |
|
|
|
|
| Muscle |
5839 |
69 |
6968 |
241 |
| Calecim |
|
|
|
|
| Skin |
9058 |
80 |
7924 |
219 |
| Melsmon |
|
|
|
|
| Muscle |
8926 |
63 |
6254 |
227 |
| Melsmon |
|
|
|
|
| Skin |
7505 |
109 |
8887 |
349 |
| Calecim+Melsmon, 4 injections/wk |
|
|
|
|
| Muscle |
5180 |
69 |
5933 |
355 |
| Calecim+Melsmon, 4 injections/wk |
|
|
|
|
| Skin |
4978 |
75 |
7511 |
300 |
| Calecim+ES, 4 injections/wk |
|
|
|
|
| Skin |
4117 |
54 |
6567 |
119 |
Table 1. Number of genes showing overlap between No injection / Young mouse changes and No injection / Treatment changes in different tissues. All experiments combined
Cluster analysis
Cluster analysis shows simultaneous comparison of 7 different samples: old and young controls, as well Melsmon, Calecim, ES cell treatments, both once per week and 4 times per week (Figure 5 and 6). Melsmon had the best effect on muscle while Calecim produced the best results with skin – all in terms of producing the expression profiles closer to that of the young mouse.
Treated mice have different expression profiles than the old untreated ones. And it was true for all treatments and samples. these results indicate that the treatments cause changes.
Old untreated mice and young mice are more similar than the treated mice. What we can conclude is that we probably overstimulated the tissues so that they produced stronger changes in gene expression than even what we normally see in younger mice. And this is probably true because younger mice produce growth factors naturally, and in treated mice we overloaded them with those factors, so the changes might have been more robust. Treated and young profiles are only similar for Melsmon muscle; this was never found for Melsmon skin or any other treatment.
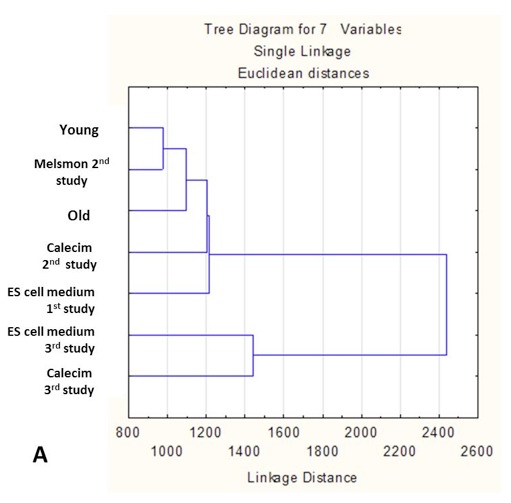
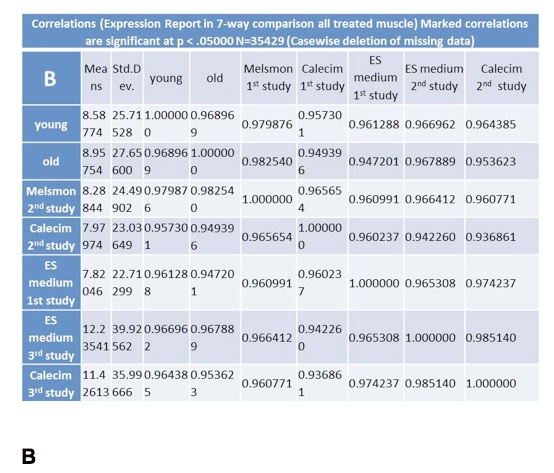
Figure 5. Microarray analysis of gene expression in muscle: Cluster analysis. A. Linkage distance B. Correlations.
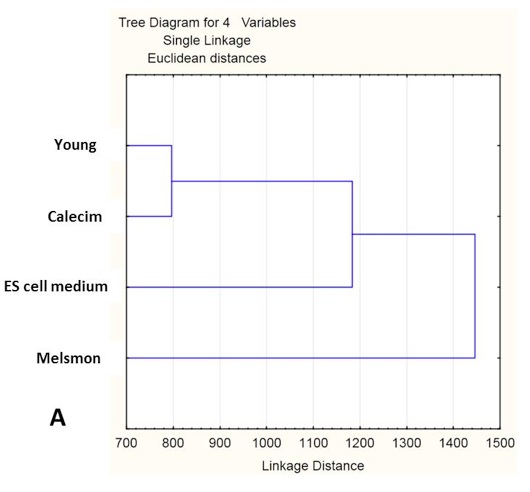
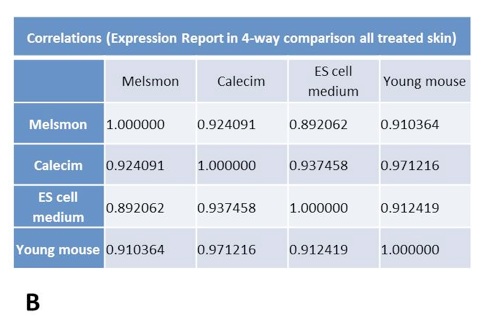
Figure 6. Microarray analysis of gene expression in skin: Cluster analysis. A. Linkage distance B. Correlations.
Discussion
Tissues from muscle, skin, heart, liver, and brain were analyzed by pathologists from Texas A&M and UCSD. Their findings indicate that all of the treated older mice in our studies did not express any kind of neoplasia or tumorgenesis. Moreover, the mice treated with one injection per week for four months likewise did not develop any kind of mutagenic response or aberrant lesions. There were many distinctive growth factors in these diverse cytokine configurations. However; they are not engendering abnormal cellular genesis. Furthermore, the mice in all the treatment groups did not develop any kind of adverse response. There were virtually no side effects with these cytokine therapies.
All the treated mice maintained a homeostatic weight as their food intake was normal. We did observe that the treated older mice were nocturnally more physically robust than the older untreated phenotypes. There were not any aberrant changes in behavior, food or water intake, as well as skewed physiological modifications in any of the mice that were injected with our distinctive cytokine configurations. Thus, the safety of these innovative peptide therapies as a result of our studies demonstrate that this prophylaxis is not problematic; and could be well tolerated by the older adult human phenotype. This supposition needs to be verified in a human trial.
We have demonstrated in culture in a previous study as well as in a mammalian model that cytokine interventions do auspiciously affect gene expression profiles to where the treated older organism becomes comparable to a younger untreated phenotype. Shane 2012 found that older human fibroblasts when treated with cytokines from embryonic stem cells expressed an overall gene profile consistent with a younger untreated phenotype. A 63 year old mother’s fibroblasts were significantly modulated to express a gene profile consistent with her 29 year old daughter as a function of this unique cytokine therapy. The results of this current study were even more dramatic in an intact mammalian organism in terms of age reversal in the treated older mice. These mice showed a gene expression profile characteristic of a younger untreated phenotype.
Single or multiple injections of stem cell therapies have not demonstrated to have any kind of ubiquitous modulation of gene expression where the mammalian organism expresses transcriptional activities characteristic of an untreated younger phenotype. There is a likelihood that subcutaneous or inner-muscular injections once a week could reverse cellular aging in the older human phenotype. Thus, our thorough review of the medical literature has not revealed any molecular strategy to ameliorate cellular gene expression to where the older organism expresses a genetic profile concordant with a younger phenotype. This novel study indicates that peptides from stem cell sources is a more efficacious strategy for reversing cellular aging as compared to any current stem cell therapy.
The research paradigm of this particular study revealed that weekly injections of cytokines were more efficacious for four months compared to a protocol of injections four times a week for 45 days. We postulate that cytokine therapies takes time to remodel or reprogram the deleterious molecular dynamics associated with cellular aging. A more rigorous study utilizing many titration regimes is needed to determine if there is an asymptotic effect that would be induced in mammalian organisms where the molecular reprogramming would gradually attenuate. However, we propose that the re-education of quiescent cellular systems as a function of these unique cytokine therapies would continue for many years where the older phenotype would be then genetically reflective of a younger organism. It is possible that four injections per week may be too excessive where this regime could induce an aberrant phenotype.
The results of this study indicate that once a week injections of a high concentration of a cytokine configuration did engender a phenotype which was more characteristic of a younger untreated organism. We likewise discern that the employment of multiple cytokine strategies were less effective than a single injection of one peptide configuration for four months. These enigmatic results likewise need to be replicated in a larger study to determine if prodigious molecular remodeling is primarily dictated by months of judicious cytokine therapy rather than a multiple weekly injection titration regime.
The skin and muscle tissue of the treated older mice expressed a gene profile consistent with a younger untreated phenotype. However, they were not analyzed by electron microscopy in terms of the histological modifications. Moreover, in our view, ostensible tissue ameliorative modifications might be observed when there is a sufficient titration period of at least a year in a human phenotype. There is some likelihood that electron microscopy would reveal propitious structural changes in certain tissues as a function of a more youthful protein expression, as a result of a mitigation of skewed protein secretion. The structural dynamics of a tissue matrix of an older phenotype is influenced by compromised protein expression which is governed by age related mollified transcriptional processes.
All the cytokine therapies utilized in this study influenced thousands of genes to be up or down regulated as shown in the tables of the results section. Moreover, many genes were highly impacted by these cytokine matrixes where there were at least a two fold up and down regulation. Cytokine matrixes from stem cells derived from human umbilical cord lining demonstrated the most profound effect in terms of reversing aging in the older mouse phenotype. The molecular phenomena of mammalian aging involves much more than up and down regulation of gene expression; however it is possible to modulate cellular transcription and translation which would then engender inordinate physiological remodeling.
In the experiment employing cytokines from embryonic stem cells the older treated mouse demonstrated a gene profile 60 percent similar to the younger untreated organism. Conversely, the treated older mice were only 40 percent similar to the older untreated mice. In a second experiment, peptide matrixes from umbilical cord lining stem cells induced older treated mice to become in terms of their gene expression 70 percent similar to the younger untreated phenotype. It was observed that the older treated mice as well as the younger untreated organisms were only 28 percent similar to the older untreated mice. Peptides from placenta extract induced a gene expression profile consistent with cytokines derived from the stem cells of umbilical cord lining.
Treated older mice in all the experiments became at least 70 percent dissimilar in terms of their gene expression profile to the older untreated mice. Thus, they were much more analogous in terms of their genetic behavior to the younger untreated phenotype as compared to the older controls. We will be undertaking another research study to further support the findings of this proof of concept study. Our overseas collaborators are commencing human studies to determine what are the actual physiological effects in terms of tissue translation of these cytokine therapies. It is evident that the biological modulation of cellular behavior through cytokine therapies are much more effective than any existing stem cell therapies for reversing aging. These cytokine therapies are not inducing any kind of pathologies; and could possibly be used in a medicinal manner to treat certain morbidities.
This innovative study represents a paradigm shift in regenerative medicine. Human studies are needed to comprehend the degree of translational modulation of compromised tissues which have been impacted by molecular aging. This current study as well as an earlier human fibroblast research project indicate that quiescent cellular machinations can be enhanced and more youthfully optimized as a function of peptide configuration therapies derived from stem cell sources. Our future research projects will more fully comprehend how cytokine matrixes engenders protein expression changes, and how this translates into the optimal amelioration of tissue dynamics in an older mammalian phenotype.
References
- Rando TA. Stem Cells, Aging, and guest for immortality, Nature 2006.
- Carlson MA, Conboy IM regulations the notch pathway in embryonic, adult and old stems.
- Sashs DL, Voorhess, JJ 2011 Age Reversing and Devices in Dermatology Clin Pharmacol Ther
- Fuller, J Hematopoietic Stem Cells and Aging. SCl. Aging Knowledge Environ
- Clifford DM, Fisher SA, Brunskill SJ, Doree C, Mathur A, Watt S, Martin Rendon E,* Stem cell treatment for acute myocardial infarction, The Cochrane Review, February, 2012.
- Beitnes JO, Oie E, Shahdadfar A, Karlsen T, Muller RM, Aakhys S, Reinholt FP, Brinchmann JE. INtramyocardial injections of human mesenchymal stem cells following acute myocardial infarcation modulate scar formation and improve left ventricular function. Cell Transplantation, March 8, 2012
- Rodrigo S, van Ramshorst V, Beeres S, Younis IA Dibbets-Schneider P, Fibbe W, Zwaginga JJ, van der Wall E, Schalij M, Bax J, Atsma D- Intramyocardial Injection of Bone Marrow Derived, Ex Vivo Expanded, Mesenchymal Stem Cells in Acute Myocardial Infarction Patients is Safe and Improves Myocardial Perfusion at 3 years follow-up Coll Cardiol.
- Shane R, Crain A, Golovko A- Propitious Modulation: Fibroblast Quiescence Mediated by Cytokine Expression from Embryonic Stem Cells; Age Management Medicine E-Journal, October 2012
- Clifford DM et al op. cit., 2012
- Beitnes JO et al op. cit., 2012
- Conboy IM and Rando TA- Heterochronic parabiosis for the study of the effects of aging on stem cells and their niches; Cell Cycle
- Conboy IM, Conboy MJ, Wagers AJ, Girma ER, Weissman IL & Rando TA; Rejuvenation of aged progenitor cells by exposure to a young systemic environment; Nature
Ron Shane, Ph.D., OMD, is currently a research scholar at U.C. San Diego, working in the field of neuroendocrinology and optimal physiology. Contact Dr. Shane at zenbeautyinstitute@gmail.com or drron73@gmail.com.
Return to June 2013 e-Journal 